The tarot cards are the most famous and most mysterious of all known ...

A molecular biologist is a medical researcher whose mission is, no less, to save humanity from dangerous diseases. Among such diseases, for example, oncology, which today has become one of the main causes of death in the world, is only slightly behind the leader - cardiovascular diseases. New methods of early diagnosis of oncology, prevention and treatment of cancer are a priority task of modern medicine. Molecular oncology biologists develop antibodies and recombinant (genetically engineered) proteins for early diagnosis or targeted drug delivery in the body. Specialists in this field use the most modern achievements of science and technology to create new organisms and organic substances with the aim of their further use in research and clinical activities. Among the methods used by molecular biologists are cloning, transfection, infection, polymerase chain reaction, gene sequencing, and others. One of the companies interested in molecular biologists in Russia is PrimeBioMed LLC. The organization is engaged in the production of antibodies reagents for the diagnosis of cancer. Such antibodies are mainly used to determine the type of tumor, its origin and malignancy, that is, the ability to metastasize (spread to other parts of the body). Antibodies are applied to thin sections of the examined tissue, after which they bind in cells with certain proteins - markers that are present in tumor cells, but absent in healthy cells and vice versa. Further treatment is prescribed depending on the results of the study. Among the clients of "PrimeBioMed" there are not only medical, but also scientific institutions, since antibodies can also be used to solve research problems. In such cases, unique antibodies can be produced that can bind to the protein under study, for a specific task on a special order. Another promising area of \u200b\u200bthe company's research is the targeted (targeted) delivery of drugs in the body. In this case, antibodies are used as transport: with their help, drugs are delivered directly to the affected organs. Thus, the treatment becomes more effective and has fewer negative consequences for the body than, for example, chemotherapy, which affects not only cancer cells, but also other cells. The profession of a molecular biologist is expected to become more and more in demand in the coming decades: with an increase in the average life expectancy of a person, the number of oncological diseases will increase. Early diagnosis of tumors and innovative treatments using substances obtained by molecular biologists will save lives and improve its quality for a huge number of people.
Molecular biology / m ə lɛ toJʊ lər / is a branch of biology that deals with the molecular basis of biological activity between biomolecules in various cell systems, including interactions between DNA, RNA, proteins and their biosynthesis, as well as the regulation of these interactions. Recording in nature in 1961, Astbury described molecular biology:
Not so much a technique as an approach, an approach from the point of view of the so-called fundamental sciences with the leading idea of \u200b\u200bsearching below the large-scale manifestations of classical biology for the corresponding molecular plan. It is concerned, in particular, with forms biological molecules and [...] predominantly three-dimensional and structural - which does not mean, however, that this is just a refinement of morphology. He must at the same time investigate genesis and function.
Researchers in the field of molecular biology use specific methods of growing molecular biology, but more and more combine them with methods and ideas from genetics and biochemistry. There is no definite line between these disciplines. This is illustrated in the following diagram, which depicts one possible kind of relationship between fields:
One of the most basic molecular biology techniques for studying protein function is molecular cloning. In this technique, DNA encoding a protein of interest is cloned by polymerase chain reaction (PCR) and / or restriction enzymes in a plasmid (expression vector). The vector has 3 distinctive features: an origin of replication, a multiple cloning site (MCS), and a selectable marker, usually antibiotic resistant. The upstream multiple cloning sites are the promoter regions and transcriptional initiation sites that regulate the expression of the cloned gene. This plasmid can be inserted into either bacterial or animal cells. The introduction of DNA into bacterial cells can be done by transformation using naked DNA uptake, conjugation using cell-cell contacts, or by transduction using a viral vector. The introduction of DNA into eukaryotic cells, such as animal cells, by physical or chemical means, is called transfection. Several different transfection methods are available such as calcium phosphate transfection, electroporation, microinjection, and liposomal transfection. The plasmid can be integrated into the genome, resulting in stable transfection, or it can remain independent of the genome, called transient transfection.
DNA encoding proteins of interest are now inside the cell, and proteins can now be expressed. Diverse systems, such as inducible promoters and specific cellular signaling factors, help express protein interest at high levels. Large amounts of protein can then be recovered from the bacterial or eukaryotic cell. The protein can be tested for enzymatic activity in various situations, the protein can be crystallized so its tertiary structure can be studied, or, in the pharmaceutical industry, the activity of new drugs against the protein can be studied.
Terms northern , west and oriental blotting gets from what was originally molecular biology a joke that played on the term Southernnet , following the procedure described by Edwin Southern for BLOTTED DNA hybridization. Patricia Thomas, developer of RNA blotting, which then became known as north - blotting , don't really use that term.
Named after its inventor, biologist Edwin South, the Southern blot is a method for testing for the presence of a specific DNA sequence in a DNA sample. DNA samples before or after the restriction enzyme (restriction enzyme) digestions are separated by gel electrophoresis and then transferred to the membrane by blotting by capillary action. The membrane is then exposed to a labeled DNA probe that has a base sequence complementary to that on the DNA of interest. Southern blotting is less widely used in the scientific laboratory due to the ability of other methods, such as PCR, to detect specific DNA sequences from DNA samples. These blots are still used for some applications, however, such as measuring a transgene copy number in transgenic mice or in gene engineering of knockout embryonic stem cell lines.
Northern blot chart
Clinical research and medical therapies arising from molecular biology are partially covered by gene therapy. The application of molecular biology or molecular cell biology approaches to medicine is now called molecular medicine. Molecular biology also plays an important role in understanding the formation, actions and regulations of various parts of cells, which can be used to effectively target new drugs, diagnose disease, and understand cell physiology.
Molecular biology, a science that sets as its task the cognition of the nature of the phenomena of life by studying biological objects and systems at a level approaching the molecular level, and in some cases even reaching this limit. The ultimate goal in this case is to find out how and to what extent the characteristic manifestations of life, such as heredity, reproduction of one's own kind, protein biosynthesis, excitability, growth and development, storage and transmission of information, energy conversion, mobility, etc. , are due to the structure, properties and interaction of molecules of biologically important substances, primarily two main classes of high molecular weight biopolymers - proteins and nucleic acids. A distinctive feature of M. b. - study of the phenomena of life on inanimate objects or those that are inherent in the most primitive manifestations of life. These are biological formations from the cellular level and below: subcellular organelles, such as isolated cell nuclei, mitochondria, ribosomes, chromosomes, cell membranes; further - systems that stand on the border of living and inanimate nature - viruses, including bacteriophages, and ending with molecules of the most important components of living matter - nucleic acids and proteins.
The foundation on which M. b. Developed was laid by such sciences as genetics, biochemistry, physiology of elementary processes, and so on. According to the origins of his development, M. b. inextricably linked to molecular genetics, which continues to be an important part of
A distinctive feature of M. b. is its three-dimensionality. M.'s essence. M. Perutz sees it in interpreting biological functions in terms of molecular structure. M. b. aims to get answers to the question "how", having learned the essence of the role and participation of the entire structure of the molecule, and to the questions "why" and "why", having clarified, on the one hand, the relationship between the properties of the molecule (again, primarily proteins and nucleic acids) and the functions it performs and, on the other hand, the role of such individual functions in the general complex of manifestations of vital activity.
Major advances in molecular biology. Here is a far from complete list of these achievements: disclosure of the structure and mechanism of the biological function of DNA, all types of RNA and ribosomes, disclosure of the genetic code; discovery of reverse transcription, i.e. DNA synthesis on an RNA template; study of the mechanisms of the functioning of respiratory pigments; discovery of three-dimensional structure and its functional role in the action of enzymes, the principle of matrix synthesis and mechanisms of protein biosynthesis; disclosure of the structure of viruses and mechanisms of their replication, primary and, partially, the spatial structure of antibodies; isolation of individual genes, chemical and then biological (enzymatic) synthesis of a gene, including human, outside the cell (in vitro); transfer of genes from one organism to another, including human cells; the rapidly advancing deciphering of the chemical structure of an increasing number of individual proteins, mainly enzymes, as well as nucleic acids; detection of the phenomena of "self-assembly" of some biological objects of increasing complexity, starting from nucleic acid molecules and moving to multicomponent enzymes, viruses, ribosomes, etc .; elucidation of allosteric and other basic principles of regulation biological functions and processes.
The problems of molecular biology. Along with the indicated important tasks of M. b. (cognition of the patterns of "recognition", self-assembly and integration) an urgent direction of scientific search for the near future is the development of methods that make it possible to decipher the structure, and then the three-dimensional, spatial organization of high molecular weight nucleic acids. All the most important methods, the use of which ensured the emergence and success of medical science, were proposed and developed by physicists (ultracentrifugation, X-ray structural analysis, electron microscopy, nuclear magnetic resonance, etc.). Almost all new physical experimental approaches (for example, the use of computers, synchrotron, or bremsstrahlung, radiation, laser technology, etc.) open up new possibilities for an in-depth study of the problems of medical science. Among the most important tasks of a practical nature, the answer to which is expected from M. b., In the first place is the problem of the molecular basis of malignant growth, then - the ways of preventing, and perhaps overcoming, hereditary diseases - "molecular diseases". Elucidation of the molecular basis of biological catalysis, that is, the action of enzymes, will be of great importance. Among the most important modern directions of M. b. should include the desire to decipher the molecular mechanisms of action of hormones, toxic and medicinal substances, as well as to clarify the details of the molecular structure and functioning of such cellular structures as biological membranes involved in the regulation of the processes of penetration and transport of substances. More distant goals of M. b. - cognition of the nature of nervous processes, memory mechanisms, etc. One of the important emerging sections of M. b. - the so-called. genetic engineering, which sets as its task the purposeful operation of the genetic apparatus (genome) of living organisms, starting with microbes and lower (unicellular) ones and ending with humans (in the latter case, primarily for the purpose of radical treatment of hereditary diseases and correction of genetic defects).
The most important areas of MB:
- Molecular genetics - the study of the structural and functional organization of the genetic apparatus of the cell and the mechanism of realization of hereditary information
- Molecular Virology - the study of the molecular mechanisms of interaction of viruses with cells
- Molecular immunology - the study of the patterns of the body's immune reactions
- Molecular developmental biology - the study of the appearance of different qualities of cells in the course of individual development of organisms and specialization of cells
The main objects of research: Viruses (including bacteriophages), Cells and subcellular structures, Macromolecules, Multicellular organisms.
Advances in the study of nucleic acids and protein biosynthesis have led to the creation of a number of methods that are of great applied importance in medicine, agriculture, and a number of other industries.
After the genetic code and the basic principles of storing and realizing hereditary information were studied, the development of molecular biology came to a dead end, since there were no methods that allowed to manipulate genes, isolate and change them. The emergence of these methods occurred in the 1970s and 1980s. This gave a powerful impetus to the development of this field of science, which is still flourishing today. First of all, these methods concern the production of individual genes and their introduction into cells of other organisms (molecular cloning and transgenesis, PCR), as well as methods for determining the sequence of nucleotides in genes (DNA and RNA sequencing). These methods will be discussed in more detail below. We will start with the simplest basic method, electrophoresis, and then move on to more complex methods.
This is the basic method of working with DNA, used in conjunction with almost all other methods to isolate the desired molecules and analyze the results. To separate DNA fragments by length, gel electrophoresis is used. DNA is an acid, its molecules contain residues of phosphoric acid, which split off a proton and acquire a negative charge (Fig. 1).
Therefore, in an electric field, DNA molecules move to the anode - a positively charged electrode. This occurs in an electrolyte solution containing charge carrier ions, so that this solution conducts current. To separate the fragments, a dense polymer gel (agarose or polyacrylamide) is used. DNA molecules become entangled in it the more, the longer they are, and therefore the longest molecules move the slowest, and the shortest ones - the fastest (Fig. 2). Before or after electrophoresis, the gel is treated with dyes that bind to DNA and fluoresce in ultraviolet light, and a picture of bands in the gel is obtained (see Fig. 3). To determine the lengths of the DNA fragments of the sample, they are compared with a marker - a set of fragments of standard lengths applied in parallel on the same gel (Fig. 4).



The most important tools for working with DNA are enzymes that transform DNA in living cells: DNA polymerases, DNA ligases, and restriction endonucleases, or restriction endonucleases. DNA polymerase carry out matrix DNA synthesis, which allows DNA to be multiplied in a test tube. DNA ligases sew together DNA molecules or heal gaps in them. Restriction endonucleases, or restriction enzymes, cut DNA molecules in strictly defined sequences, which allows you to cut out individual fragments from the total mass of DNA. These fragments may in some cases contain separate genes.
The sequences recognized by restriction endonucleases are symmetrical, and breaks can occur in the middle of such a sequence or with a shift (at the same place in both DNA strands). Action scheme different types restriction enzymes are shown in Fig. 1. In the first case, so-called "blunt" ends are obtained, and in the second, "sticky" ends. In the case of "sticky" ends of the bottom, the chain turns out to be shorter than the other, a single-stranded section is formed with a symmetrical sequence identical at both ends formed.
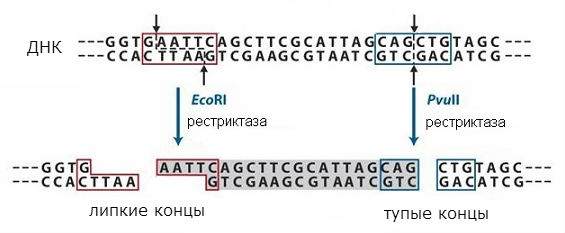
The terminal sequences will be the same when any DNA is cleaved with a given restriction enzyme, and can be reattached since they have complementary sequences. They can be stitched together using DNA ligase and get a single molecule. Thus, it is possible to combine fragments of two different DNA and obtain the so-called recombinant DNA... This approach is used in the method of molecular cloning, which allows you to obtain individual genes and introduce them into cells, which can form a protein encoded in the gene.
Molecular cloning uses two DNA molecules - an insert containing the gene of interest, and vector - DNA serving as a carrier. The insert is "sewn" into the vector using enzymes to obtain a new, recombinant DNA molecule, then this molecule is introduced into host cells, and these cells form colonies on a nutrient medium. A colony is the offspring of one cell, that is, a clone, all cells in a colony are genetically identical and contain the same recombinant DNA. Hence the term "molecular cloning", that is, obtaining a clone of cells containing the DNA fragment of interest. After the colonies containing the insert of interest to us are obtained, it is possible to characterize this insert by various methods, for example, to determine its exact sequence. Cells can also produce an insert-encoded protein if it contains a functional gene.
When a recombinant molecule is introduced into cells, genetic transformation of these cells occurs. Transformation - the process of absorption by a cell of an organism of a free DNA molecule from the environment and its incorporation into the genome, which leads to the appearance in such a cell of new inherited traits characteristic of a DNA donor organism. For example, if the inserted molecule contains the gene for resistance to the antibiotic ampicillin, then the transformed bacteria will grow in its presence. Before transformation, ampicillin caused their death, that is, a new trait appears in the transformed cells.
A vector must have a number of properties:
Firstly, it is a relatively small DNA molecule to be easily manipulated.
Secondly, in order for DNA to be preserved and multiplied in a cell, it must contain a specific sequence that ensures its replication (the origin of replication, or origin of replication).
Third, it must contain gene marker, which ensures the selection of only those cells into which the vector fell. Usually these are antibiotic resistance genes - then in the presence of an antibiotic, all cells not containing the vector die.
Gene cloning is most often carried out in bacterial cells, since they are easy to cultivate and multiply quickly. A bacterial cell usually contains one large circular DNA molecule, several million base pairs long, containing all the genes necessary for bacteria - the bacterial chromosome. In addition to it, some bacteria have small (several thousand base pairs) circular DNA called plasmids(fig. 2). They, like basic DNA, contain a sequence of nucleotides that ensure the ability of DNA to replicate (ori). Plasmids replicate independently of the main (chromosomal) DNA, therefore they are present in the cell in a large number of copies. Many of these plasmids carry antibiotic resistance genes that distinguish plasmid-bearing cells from normal cells. Plasmids carrying two genes are used more often, providing resistance to two antibiotics, for example, to tetracycline and amicillin. Exists simple methods isolation of such plasmid DNA, free from DNA of the main chromosome of the bacterium.

The transfer of genes from one organism to another is called transgenesis, and such modified organisms - transgenic... By transferring genes into the cells of microorganisms, recombinant protein preparations are obtained for the needs of medicine, in particular, human proteins that do not cause immune rejection - interferons, insulin and other protein hormones, cellular growth factors, as well as proteins for the production of vaccines. In more complex cases, when the modification of proteins proceeds correctly only in eukaryotic cells, transgenic cell cultures or transgenic animals are used, in particular, livestock (primarily goats), which secretes the necessary proteins into milk, or proteins are isolated from their blood. This is how antibodies, blood clotting factors and other proteins are obtained. By the method of transgenesis, crop plants are obtained that are resistant to herbicides and pests and have other useful properties... With the help of transgenic microorganisms, they purify wastewater and fight pollution, there are even transgenic microbes that can break down oil. In addition, transgenic technologies are indispensable in scientific research - the development of biology today is unthinkable without the routine use of methods of modification and gene transfer.
molecular cloning technology
To obtain an individual gene from any organism, all chromosomal DNA is isolated from it and cleaved with one or two restriction enzymes. Enzymes are selected so that they do not cut the gene of interest to us, but make breaks along its edges, and make 1 break in the plasmid DNA in one of the resistance genes, for example, to ampicillin.
The molecular cloning process includes the following steps:
Cutting and stitching - construction of a single recombinant molecule from an insert and a vector.
Transformation is the introduction of a recombinant molecule into cells.
Selection - selection of cells that have received an insert vector.
Plasmid DNA is treated with the same restriction enzymes, and it turns into a linear molecule if such a restriction enzyme is selected that introduces 1 gap into the plasmid. As a result, the ends of all the resulting DNA fragments end up with the same sticky ends. When the temperature is lowered, these ends are connected randomly, and they are ligated with DNA ligase (see Fig. 3).

Get a mixture of circular DNA different composition: some of them will contain a specific DNA sequence of chromosomal DNA, linked to bacterial DNA, others - fragments of chromosomal DNA linked together, and still others - a reduced circular plasmid or its dimer (Fig. 4).

Then this mixture is carried out genetic transformation bacteria that do not contain plasmids. Transformation - the process of absorption by a cell of an organism of a free DNA molecule from the environment and its incorporation into the genome, which leads to the appearance in such a cell of new inherited traits characteristic of a DNA donor organism. Only one plasmid can enter and multiply in each cell. Such cells are placed on a solid nutrient medium containing the antibiotic tetracycline. The cells that did not get the plasmid will not grow on this medium, and the cells carrying the plasmid form colonies, each of which contains the descendants of only one cell, i.e. all cells in a colony carry the same plasmid (see Fig. 5).

Next, the task is to select only the cells into which the vector with the insertion fell, and to distinguish them from the cells that carry only the vector without the insertion or do not carry the vector at all. This process of selecting the desired cells is called breeding... For this, use selective markers - usually antibiotic resistance genes in the vector, and selective mediacontaining antibiotics or other substances that provide selection.
In our example, cells from colonies grown in the presence of ampicillin are subcultured into two media: the first contains ampicillin, and the second contains tetracycline. Colonies containing only the plasmid will grow on both media, while colonies with inserted chromosomal DNA in the plasmids will not grow on the medium with tetracycline (Fig. 5). Among them, by special methods, those that contain the gene of interest to us are selected, grown in sufficient quantities and plasmid DNA is isolated. The individual gene of interest is excised from it using the same restriction enzymes that were used to obtain recombinant DNA. The DNA of this gene can be used to determine the sequence of nucleotides, introduce it into an organism to obtain new properties, or synthesize the desired protein. This method of isolating genes is called molecular cloning.
It is very convenient to use fluorescent proteins as marker genes in studies of eukaryotic organisms. Gene of the first fluorescent protein, green fluorescent protein (GFP) was isolated from the jellyfish Aqeuorea victoria and introduced into various model organisms (see Fig. 6) In 2008, O. Simomura, M. Chalfi and R. Tsien received the Nobel Prize for the discovery and use of this protein.

Then the genes of other fluorescent proteins were isolated - red, blue, yellow. These genes have been artificially modified to produce proteins with the desired properties. The variety of fluorescent proteins is shown in Fig. 7, which shows a Petri dish with bacteria containing genes for various fluorescent proteins.

use of fluorescent proteins
The fluorescent protein gene can be ligated with the gene of any other protein, then a single protein will be formed during translation - a translationally fusion protein, or fusion (fusion protein) that fluoresces. Thus, you can study, for example, the localization (location) of any proteins of interest in the cell, their movement. By expressing fluorescent proteins only in certain types of cells, it is possible to mark cells of these types in a multicellular organism (see Fig. 8 - the mouse brain, in which individual neurons have different colors due to a certain combination of genes of fluorescent proteins). Fluorescent proteins are an indispensable tool in modern molecular biology.

Another method of obtaining genes is called polymerase chain reaction (PCR)... It is based on the ability of DNA polymerases to complete the second strand of DNA along the complementary strand, as it happens in cells during DNA replication.
The origin of replication in this method is defined by two small pieces of DNA called seeds,or primers... These primers are complementary to the ends of the gene of interest on two DNA strands. First, the chromosomal DNA, from which the gene must be isolated, is mixed with seeds and heated to 99 ° C. This leads to the rupture of hydrogen bonds and the divergence of DNA strands. After that, the temperature is lowered to 50-70 ° C (depending on the length and sequence of seeds). Under these conditions, the primers are attached to the complementary regions of chromosomal DNA, forming a regular double helix (see Fig. 9). After that, a mixture of all four nucleotides required for DNA synthesis and DNA polymerase are added. The enzyme lengthens the primers by building double-stranded DNA from where the primers are attached, i.e. from the ends of the gene to the end of the single-stranded chromosomal molecule. 
If the mixture is now heated again, the chromosomal and newly synthesized chains will disperse. After cooling, seeds will again join them, which are taken in large excess (see Fig. 10).

On the newly synthesized chains, they will attach not to the end from which the first synthesis began, but to the opposite end, since the DNA chains are antiparallel. Therefore, in the second cycle of synthesis, only the sequence corresponding to the gene will be completed on such chains (see Fig. 11).
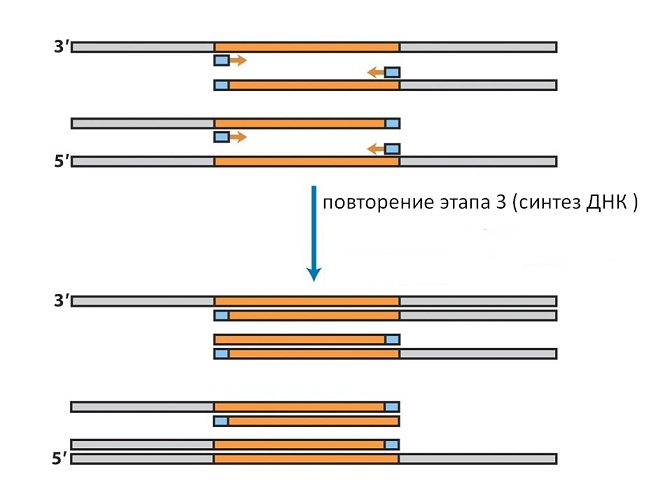
In this method, DNA polymerase from thermophilic bacteria is used, which can withstand boiling and operates at temperatures of 70-80 ° C, it does not need to be added every time, but it is enough to add it at the beginning of the experiment. By repeating the heating and cooling procedures in the same sequence, we can double in each cycle the number of sequences bounded at both ends by the introduced seeds (see Fig. 12).

After about 25 such cycles, the copy number of the gene will increase more than a million times. Such quantities can be easily separated from the chromosomal DNA introduced into the test tube and used for various purposes.
Another important achievement is the development of methods for determining the sequence of nucleotides in DNA - dNA sequencing(from the English sequence - sequence). To do this, it is necessary to obtain genes pure from other DNA by one of the described methods. Then the DNA strands are separated by heating and a primer labeled with radioactive phosphorus or a fluorescent label is added to them. Note that one seed is taken, complementary to one strand. Then DNA polymerase and a mixture of 4 nucleotides are added. Such a mixture is divided into 4 parts, and to each one of the nucleotides is added, modified so that it does not contain a hydroxyl group at the third atom of deoxyribose. If such a nucleotide is included in the synthesized DNA strand, then its elongation cannot continue, because the polymerase will have nowhere to attach the next nucleotide. Therefore, DNA synthesis is terminated after the inclusion of such a nucleotide. Much less of such nucleotides, called dideoxynucleotides, are added than ordinary ones, so chain termination occurs only occasionally and in each chain in different places. The result is a mixture of chains of different lengths, each with the same nucleotide at the end. Thus, the chain length corresponds to the nucleotide number in the studied sequence, for example, if we had an adenyl dideoxynucleotide, and the resulting chains were 2, 7, and 12 nucleotides long, then there was adenine in the gene in the second, seventh and twelfth positions. The resulting mixture of chains can be easily separated by size using electrophoresis, and the synthesized chains can be identified by radioactivity on an X-ray film (see Fig. 10).

It turns out the picture shown at the bottom of the figure, called a radio autograph. Moving along it from bottom to top and reading the letter above the columns of each zone, we get the nucleotide sequence shown in the figure to the right of the autograph. It turned out that the synthesis is stopped not only by dideoxynucleotides, but also by nucleotides, in which some chemical group, for example, a fluorescent dye, is attached to the third position of the sugar. If each nucleotide is marked with its own dye, then the zones obtained during the separation of the synthesized strands will glow with different light. This makes it possible to carry out the reaction in one test tube simultaneously for all nucleotides and dividing the obtained chains by length, to identify nucleotides by color (see Fig. 11).

Such methods made it possible to determine the sequence of not only individual genes, but also to read entire genomes. Even faster methods of determining the sequences of nucleotides in genes have now been developed. If the pair human genome was deciphered by a large international consortium using the first given method in 12 years, the second, using the second, in three years, now this can be done in a month. This allows predicting a person's predisposition to many diseases and taking measures in advance to avoid them.
The development of biochemistry, biophysics, genetics, cytochemistry, many branches of microbiology and virology around the beginning of the 40s of the XX century. brought close to the study of life phenomena at the molecular level. The successes achieved by these sciences, simultaneously and from different sides, led to the realization of the fact that it is at the molecular level that the main control systems of the body function and that further progress of these sciences will depend on the disclosure of the biological functions of molecules that make up the body of organisms, their participation in synthesis and decay, mutual transformations and reproduction of compounds in the cell, as well as the exchange of energy and information that occurs during this. So, at the junction of these biological disciplines with chemistry and physics, a completely new branch arose - molecular biology.
Unlike biochemistry, the attention of modern molecular biology is focused mainly on the study of the structure and function of the most important classes of biopolymers - proteins and nucleic acids, the former of which determine the very possibility of metabolic reactions, and the latter - the biosynthesis of specific proteins. Therefore, it is clear that it is impossible to draw a clear distinction between molecular biology and biochemistry, the corresponding sections of genetics, microbiology and virology.
The emergence of molecular biology was closely related to the development of new research methods, which have already been discussed in the relevant chapters. Along with the development of electron microscopy and other methods of microscopic technology, the methods of fractionation of cellular elements, developed in the 50s, played an important role. They were based on improved methods of differential centrifugation (A. Claude, 1954). By this time, there were already fairly reliable methods for the isolation and fractionation of biopolymers. This includes, in particular, the method proposed by A. Tiselius (1937; Nobel Prize, 1948) for fractionation of proteins using electrophoresis, methods for the isolation and purification of nucleic acids (E. Key, A. Downs, M. Sevag, A. Mirsky, etc. ). In parallel, various methods of chromatographic analysis were developed in many laboratories around the world (A. Martin and R. Sing, 1941; Nobel Prize, 1952), which were subsequently significantly improved.
An invaluable service in deciphering the structure of biopolymers was played by X-ray structural analysis. The basic principles of X-ray structural analysis were developed at King's College, University of London under the leadership of W. Bragg by a group of researchers, which included J. Bernal, A. Lonsdale, W. Astbury, J. Robertson and others.
Of particular note is the research of Professor Moscow state university AR Kizel on the biochemistry of protoplasm (1925 - 1929), which were of great importance for the subsequent development of molecular biology. Kizel struck a deeply rooted notion that at the heart of all protoplasm lies a special protein body - plates, which supposedly determines all of its most important structural and functional features. He showed that the plates are a protein that is found only in myxomycetes, and then at a certain stage of development, and that there is no constant component - a single skeletal protein - in the protoplasm. Thus, the study of the problem of the structure of protoplasm and the functional role of proteins took the right path and gained scope for its development. Kiesel's research has won worldwide recognition, stimulating the study of the chemistry of the constituent parts of the cell.
The term "molecular biology", first used by the English crystallographer of the University of Leeds, W. Astbury, appeared probably in the early 1940s (before 1945). The fundamental X-ray structural studies of proteins and DNA, carried out by Astbury in the 1930s, served as the basis for the subsequent successful deciphering of the secondary structure of these biopolymers. In 1963, J. Bernal wrote: "A monument to him will be erected by all molecular biology - the science that he called and actually founded" *. analysis of organic and fibrillar compounds ", published in the English journal" Nature "**. Astbury (1950) noted in his Harvey Lecture: "I am pleased that now the term molecular biology is already quite widely used, although it is unlikely that I first proposed it. I liked it and I have long tried to spread it." As early as 1950, Astbury was clear that molecular biology deals primarily with the structure and conformation of macromolecules, the study of which is crucial for understanding the functioning of living organisms.
* (Biogr. Mem. Fellows Roy. Soc, 1963, v. 9, 29.)
** (W. T. Astbury. Progress of X-ray analysis of organic and fiber structures.- Nature ,. 1946, v. 157, 121.)
*** (W. T. Astbury. Adventures in Molecular Biology... Thomas Springfield, 1952, p. 3.)
Molecular biology was and is facing, in fact, the same tasks as for all biology as a whole - the knowledge of the essence of life and its basic phenomena, in particular, such as heredity and variability. Modern molecular biology is primarily intended to decipher the structure and function of genes, the ways and mechanisms of the realization of the genetic information of organisms at different stages of ontogenesis and at different stages of its reading. It is designed to reveal the subtle mechanisms of regulation of gene activity and cell differentiation, to clarify the nature of mutagenesis and the molecular basis of the evolutionary process.
For the development of molecular biology, the following discoveries were of greatest importance. In 1944, American researchers O. Avery, C. McLeod (Nobel Prize, 1923) and M. McCarthy showed that DNA molecules isolated from pneumococci have transforming activity. After hydrolysis of these DNAs with deoxyribonuclease, their transforming activity completely disappeared. Thus, it was convincingly proved for the first time that it is DNA, not protein, that is endowed with genetic functions in a cell.
In fairness, it should be noted that the phenomenon of bacterial transformation was discovered much earlier than the discoveries of Avery, McLeod and McCarthy. In 1928, F. Griffith published an article in which he reported that after adding killed cells of an encapsulated virulent strain to non-virulent (non-encapsulated) pneumococci, the resulting mixture of cells becomes fatal to mice. Moreover, living cells of pneumococci isolated from animals infected with this mixture were already virulent and possessed a polysaccharide capsule. Thus, in this experiment, it was shown that under the influence of some components of the killed pneumococcal cells, the non-encapsulated form of bacteria is converted into a capsule-forming virulent form. 16 years later, Avery, McLeod, and McCarthy replaced killed whole pneumococcal cells in this experiment with their deoxyribonucleic acid and showed that it is DNA that has transforming activity (see also chapters 7 and 25). The significance of this discovery can hardly be overestimated. It stimulated the study of nucleic acids in many laboratories around the world and made scientists focus on DNA.
Along with the discovery of Avery, McLeod and McCarthy, by the beginning of the 50s, a fairly large amount of direct and indirect evidence had already accumulated that nucleic acids play an exclusive role in life and carry a genetic function. This, in particular, was indicated by the nature of the localization of DNA in the cell and the data of R. Vendreli (1948) that the DNA content per cell is strictly constant and correlates with the degree of ploidy: in haploid germ cells, the DNA is half that in diploid somatic cells. The genetic role of DNA was also supported by its pronounced metabolic stability. By the beginning of the 50s, a lot of various facts had accumulated, indicating that most of the known mutagenic factors act primarily on nucleic acids and, in particular, on DNA (R. Hotchkiss, 1949; G. Ephrussi-Taylor, 1951; E. Freese , 1957, etc.).
The study of various phages and viruses was of particular importance in establishing the genetic role of nucleic acids. In 1933 D. Schlesinger found DNA in the bacteriophage of E. coli. Since the isolation of W. Stanley (1935, Nobel Prize, 1946) of the tobacco mosaic virus (TMV) in the crystalline state, a new stage in the study of plant viruses began. In 1937 - 1938. F. Bowden and N. Peary, employees of the Rothamstead Agricultural Station (England), showed that many plant viruses they isolated are not globulins, but are ribonucleoproteins and contain nucleic acid as an essential component. At the very beginning of the 40s, the works of G. Schramm (1940), P.A.Agatov (1941), G. Miller and W. Stanley (1941) were published, indicating that a noticeable chemical modification of the protein component does not lead to loss of infectivity of TMV. This indicated that the protein component cannot be the carrier of the hereditary properties of the virus, as many microbiologists continued to believe. Convincing evidence in favor of the genetic role of nucleic acid (RNA) in plant viruses was obtained in 1956 by G. Schramm in Tübingen (FRG) and H. Frenkel-Konrath in California (USA). These researchers almost simultaneously and independently from each other isolated RNA from TMV and showed that it is it, and not the protein, that possesses infectivity: as a result of infection of tobacco plants with this RNA, normal viral particles were formed and multiplied in them. This meant that RNA contains information for the synthesis and assembly of all viral components, including the viral protein. In 1968, I. G. Atabekov established that protein plays an essential role in the infection of plants itself - the nature of the protein determines the spectrum of host plants.
In 1957, Frenkel-Konrat was the first to reconstruct TMV from its constituent components - RNA and protein. Along with normal particles, he obtained mixed "hybrids" in which the RNA was from one strain, and the protein from another. The heredity of such hybrids was completely determined by RNA, and the progeny of the viruses belonged to the strain whose RNA was used to obtain the original mixed particles. Later, the experiments of A. Girer, G. Schuster and G. Schramm (1958) and G. Vitman (1960 - 1966) showed that chemical modification of the TMV nucleic acid component leads to the appearance of various mutants of this virus.
In 1970 D. Baltimore and G. Temin established that the transfer of genetic information can occur not only from DNA to RNA, but also vice versa. They found in some oncogenic RNA-containing viruses (oncornaviruses) a special enzyme, the so-called reverse transcriptase, which is capable of complementarily synthesizing DNA on RNA strands. This major discovery made it possible to understand the mechanism of insertion into the host genome of the genetic information of RNA-containing viruses and to take a fresh look at the nature of their oncogenic action.
The term nucleic acid was introduced by the German biochemist R. Altmann in 1889, after these compounds were discovered in 1869 by the Swiss physician F. Miescher. Miescher extracted the pus cells with dilute hydrochloric acid for several weeks and obtained an almost pure nuclear material in the residue. He considered this material a characteristic "substance of cell nuclei and called it nuclein. In its properties, nuclein sharply differed from proteins: it was more acidic, did not contain sulfur, but it contained a lot of phosphorus, it was well soluble in alkalis, but did not dissolve in dilute acids.
Misher sent the results of his observations of the nuclein to F. Hoppe-Seiler for publication in the journal. The substance he described was so unusual (at that time, of all biological phosphorus-containing compounds, only lecithin was known) that Hoppe-Seiler did not believe Mischer's experiments, returned the manuscript to him and instructed his collaborators N. Plosh and N. Lyubavin to check his conclusions on other material ... Mischer's work "On the Chemical Composition of Pus Cells" was published two years later (1871). At the same time, the works of Hoppe-Seiler and his collaborators on the composition of pus cells, erythrocytes of birds, snakes and other cells were published. Over the next three years, nuclein was isolated from animal cells and yeast.
In his work, Misher noted that a detailed study of different nucleins can lead to the establishment of differences between them, thereby anticipating the idea of \u200b\u200bthe specificity of nucleic acids. While studying salmon milk, Miescher found that the nuclein is in the form of a salt and is associated with a basic protein, which he called protamine.
In 1879 A. Kossel began to study nucleins in the laboratory of Hoppe-Seiler. In 1881, he isolated hypoxanthine from nuclein, but at that time he still doubted the origin of this base and believed that hypoxanthine could be a product of protein degradation. In 1891, among the products of nuclein hydrolysis, Kossel discovered adenine, guanine, phosphoric acid, and another substance with sugar properties. For his research on the chemistry of nucleic acids, Kossel was awarded the Nobel Prize in 1910.
Further advances in deciphering the structure of nucleic acids are associated with the research of P. Levin and colleagues (1911 - 1934). In 1911 P. Levin and V. Jacobs identified the carbohydrate component of adenosine and guanosine; they found that D-ribose is part of these nucleosides. In 1930, Levin showed that the carbohydrate component of deoxyribonucleosides is 2-deoxy-D-ribose. From his works it became known that nucleic acids are built from nucleotides, that is, phosphorylated nucleosides. Levin believed that the main type of bond in nucleic acids (RNA) is the 2 ", 5" -phosphodiester bond. This view turned out to be wrong. Thanks to the work of the English chemist A. Todd (Nobel Prize, 1957) and his collaborators, as well as the English biochemists R. Markham and J. Smith, it became known in the early 1950s that the main type of bond in RNA is 3 ", 5" - phosphodiester bond.
Levin showed that different nucleic acids can differ in the nature of the carbohydrate component: some of them contain the sugar deoxyribose, while others contain ribose. In addition, these two types of nucleic acids differed in the nature of one of the bases: the pentose type nucleic acids contained uracil, and the deoxypentose type nucleic acids contained thymine. Deoxypentose nucleic acid (in modern terminology, deoxyribonucleic acid - DNA) was usually easily isolated in large quantities from the thymus (thymus gland) of calves. Therefore, it received the name thymonucleic acid. The source of pentose nucleic acid (RNA) was mainly yeast and wheat germ. This type has often been referred to as yeast nucleic acid.
At the beginning of the 1930s, the idea that yeast-type nucleic acid is characteristic of plant cells, and that thymonucleic acid is characteristic only of the nuclei of animal cells, took root quite firmly. The two types of nucleic acids - RNA and DNA - were called plant and animal nucleic acids, respectively. However, as shown by the early studies of A. N. Belozersky, such a division of nucleic acids is unjustified. In 1934, Belozersky first discovered thymonucleic acid in plant cells: from pea seedlings he isolated and identified thymine-pyrimidine base, which is characteristic of DNA. Then he discovered thymine in other plants (soybean seeds, beans). In 1936, A. N. Belozersky and I. I. Dubrovskaya isolated preparative DNA from horse chestnut seedlings. In addition, a series of works carried out in England in the 1940s by D. Davidson and his colleagues convincingly showed that plant nucleic acid (RNA) is contained in many animal cells.
The widespread use of the cytochemical reaction to DNA and the reaction of J. Brachet (1944) to RNA, developed by R. Felgen and G. Rosenbeck (1924), made it possible to fairly quickly and unambiguously resolve the issue of the predominant localization of these nucleic acids in the cell. It turned out that DNA is concentrated in the nucleus, while RNA is mainly concentrated in the cytoplasm. Later, it was found that RNA is contained both in the cytoplasm and in the nucleus, and in addition, cytoplasmic DNA was identified.
As for the question of the primary structure of nucleic acids, by the middle of the 40s, P. Levin's idea was firmly established in science, according to which all nucleic acids are built according to the same type and consist of the same so-called tetranucleotide blocks. Each of these blocks, according to Levin, contains four different nucleotides. The tetranucleotide theory of the structure of nucleic acids largely deprived these biopolymers of specificity. Therefore, it is not surprising that at that time all the specifics of living things were associated only with proteins, the nature of the monomers of which is much more diverse (20 amino acids).
The first breach in the theory of the tetranucleotide structure of nucleic acids was made by the analytical data of the English chemist J. Guland (1945-1947). When determining the composition of nucleic acids by nitrogen of bases, he did not receive an equimolar ratio of bases, as it should have been according to Levin's theory. Finally, the tetranucleotide theory of the structure of nucleic acids collapsed as a result of the research of E. Chargaff and his collaborators (1949 - 1951). To separate the bases that are cleaved from DNA as a result of its acid hydrolysis, Chargaff used paper chromatography. Each of these bases has been precisely determined spectrophotometrically. Chargaff noticed significant deviations from the equimolar base ratio in DNA of different origins and for the first time definitely stated that DNA has a pronounced species specificity. Thus, the hegemony of the concept of protein specificity in a living cell was ended. Analyzing DNA of different origins, Chargaff discovered and formulated unique patterns of DNA composition, which entered science under the name of Chargaff's rules. According to these rules, in all DNA, regardless of origin, the amount of adenine is equal to the amount of thymine (A \u003d T), the amount of guanine is equal to the amount of cytosine (G \u003d C), the amount of purines is equal to the amount of pyrimidines (G + A \u003d C + T), the amount bases with 6-amino groups is equal to the number of bases with 6-keto groups (A + C \u003d G + T). At the same time, despite such strict quantitative correspondences, DNA of different species differ in the value of the ratio A + T: G + C. In some DNA, the amount of guanine and cytosine prevails over the amount of adenine and thymine (Chargaff called these DNA GC-type DNA); other DNAs contained more adenine and thymine than guanine and cytosine (these DNAs were called AT-type DNA). The data obtained by Chargaff on the composition of DNA played an exceptional role in molecular biology. They formed the basis for the discovery of the structure of DNA, made in 1953 by J. Watson and F. Crick.
Back in 1938, W. Astbury and F. Bell, using X-ray structural analysis, showed that the base planes in DNA should be perpendicular to the long axis of the molecule and resemble, as it were, a stack of plates lying one above the other. As the technique of X-ray structural analysis improved by 1952 - 1953. information has accumulated that made it possible to judge the length of individual bonds and angles of inclination. This made it possible with most likely to represent the nature of the orientation of the rings of pentose residues in the sugar-phosphate backbone of the DNA molecule. In 1952 S. Farberg proposed two speculative models of DNA, which represented a single-stranded molecule folded or twisted onto itself. An equally speculative model of the structure of DNA was proposed in 1953 by L. Pauling (Nobel laureate, 1954) and R. Corey. In this model, three twisted DNA strands formed a long helix, the core of which was represented by phosphate groups, and the bases were located outside of it. By 1953 M. Wilkins and R. Franklin obtained more clear X-ray patterns of DNA. Their analysis showed the complete inconsistency of the Farberg, Pauling and Corey models. Using Chargaff's data, comparing different combinations of molecular models of individual monomers and X-ray structural analysis data, J. Watson and F. Crick in 1953 came to the conclusion that the DNA molecule must be a double-stranded helix. Chargaff's rules sharply limited the number of possible ordered base combinations in the proposed DNA model; they suggested to Watson and Crick that there must be a specific base pairing in the DNA molecule - adenine with thymine, and guanine with cytosine. In other words, adenine in one DNA chain always strictly corresponds to thymine in the other chain, and guanine in one chain necessarily corresponds to cytosine in the other. Thus, Watson and Crick were the first to formulate the principle of the complementary structure of DNA of exceptional importance, according to which one DNA strand complements another, i.e., the base sequence of one strand uniquely determines the base sequence in the other (complementary) strand. It became obvious that the very structure of DNA contains the potential for its exact reproduction. This model of the structure of DNA is now generally accepted. For deciphering the structure of DNA, Crick, Watson and Wilkins were awarded the Nobel Prize in 1962.
It should be noted that the idea of \u200b\u200ba mechanism for the exact reproduction of macromolecules and the transmission of hereditary information originated in our country. In 1927, N, K. Koltsov suggested that during cell multiplication, the reproduction of molecules occurs by exact autocatalytic reproduction of the available parent molecules. True, at that time Koltsov endowed this property not with DNA molecules, but with protein molecules, the functional significance of which was not known at that time. Nevertheless, the very idea of \u200b\u200bautocatalytic reproduction of macromolecules and the mechanism of transmission of hereditary properties turned out to be prophetic: it became the guiding idea of \u200b\u200bmodern molecular biology.
A.S. Spirin, G.N. Zaitseva, B.F. Vanyushin, S.O. Uryson, A.S. Antonov and others, carried out in the laboratory of A.N.Belozersky, A.S. diverse organisms fully confirmed the patterns discovered by Chargaff, and full compliance with the molecular model of the structure of DNA, proposed by Watson and Crick. These studies have shown that the DNA of different bacteria, fungi, algae, actinomycetes, higher plants, invertebrates and vertebrates have specific composition. Differences in the composition (content of AT-base pairs) are especially pronounced in microorganisms, being an important taxonomic feature. In higher plants and animals, species variations in the composition of DNA are much less pronounced. But this does not mean at all that their DNA is less specific. In addition to the composition of the bases, the specificity is largely determined by their sequence in the DNA strands.
Along with the usual bases, additional nitrogenous bases were found in DNA and RNA. Thus, G. White (1950) found 5-methylcytosine in the DNA of plants and animals, and D. Dunn and J. Smith (1958) found methylated adenine in some DNA. For a long time methylcytosine was considered a hallmark of the genetic material of higher organisms. In 1968 A. N. Belozersky, B. F. Vanyushin, and N. A. Kokurina established that it can also be found in the DNA of bacteria.
In 1964, M. Gold and J. Hurwitz discovered a new class of enzymes that naturally modify DNA — its methylation. After this discovery, it became clear that minor (contained in small amounts) bases appear already on the finished polynucleotide DNA chain as a result of specific methylation of cytosine and adenine residues in special sequences. In particular, according to BF Vanyushin, Ya. I. Buryanov and AN Belozerskiy (1969) methylation of adenine in E. coli DNA can occur in termination codons. According to A.N.Belozersky and colleagues (1968 - 1970), as well as M. Meselson (USA) and V. Arber (Switzerland) (1965 - 1969), methylation gives DNA molecules unique individual features and, in combination with the action of specific nucleases, is part of a complex mechanism that controls the synthesis of DNA in the cell. In other words, the nature of methylation of a particular DNA determines the question of whether it can multiply in a given cell.
At almost the same time, the isolation and intensive study of DNA methylases and restriction endonucleases began; in 1969 - 1975 established nucleotide sequences recognized in DNA by some of these enzymes (H. Boyer, H. Smith, S. Lynn, K. Murray). When different DNAs are hydrolyzed with a restriction enzyme, rather large fragments with the same sticky ends are cleaved. This makes it possible not only to analyze the structure of genes, as is done in small viruses (D. Nathans, S. Adler, 1973 - 1975), but also to construct various genomes. With the discovery of these specific restriction enzymes, genetic engineering has become a tangible reality. Genes of different origins inserted into small plasmid DNAs are already easily introduced into different cells. Thus, a new type of biologically active plasmid was obtained, which gives resistance to some antibiotics (S. Cohen, 1973), the ribosomal genes of frog and Drosophila were introduced into the plasmids of E. coli (J. Morrow, 1974; H. Boyer, D. Hogness, R. Davis , 1974 - 1975). Thus, real ways have been opened for obtaining fundamentally new organisms by introducing and integrating various genes into their gene pool. This discovery can be directed to the benefit of all mankind.
In 1952 G. White and S. Cohen discovered that the DNA of T-even phages contains an unusual base - 5-hydroxymethylcytosine. Later, from the works of E. Vol'kin and R. Sinsheimer (1954) and Cohen (1956), it became known that oxymethylcytosine residues can be fully or partially glucosidized, as a result of which the phage DNA molecule is protected from the hydrolytic action of nucleases.
In the early 50s from the works of D. Dunn and J. Smith (England), S. Zamenhof (USA) and A. Wacker (Germany), it became known that many artificial analogs of bases can be included in DNA, sometimes replacing up to 50% thymine. Typically, these substitutions lead to errors in replication, DNA transcription and translation and to the emergence of mutants. Thus, J. Marmur (1962) established that the DNA of some phages contains oxymethyluracil instead of thymine. In 1963 I. Takahashi and J. Marmur discovered that the DNA of one of the phages contains uracil instead of thymine. Thus, another principle by which nucleic acids were previously separated has collapsed. Since the time of P. Levin's work, it was believed that the hallmark of DNA is thymine, and RNA is uracil. It became clear that this feature is not always reliable, and the fundamental difference in the chemical nature of the two types of nucleic acids, as it seems to date, is only the nature of the carbohydrate component.
In the study of phages, many unusual signs of the organization of nucleic acids were discovered. Since 1953, it has been believed that all DNA is double-stranded linear molecules, while RNA is only single-stranded. This situation was significantly shaken in 1961, when R. Sinsheimer discovered that the DNA of the phage φ X 174 is represented by a single-stranded circular molecule. True, later it turned out that in this form this DNA exists only in the vegetative phage particle, and the replicative form of this phage's DNA is also double-stranded. In addition, it turned out quite unexpectedly that the RNA of some viruses can be double stranded. This new type of macromolecular organization of RNA was discovered in 1962 by P. Gomatos, I. Tamm and other researchers in some animal viruses and in the wound tumor virus of plants. Recently, V. I. Agol and A. A. Bogdanov (1970) established that in addition to linear RNA molecules, there are also closed or cyclic molecules. Cyclic double stranded RNA was detected by them, in particular, in the encephalomyelocarditis virus. Thanks to the works of H. Devo, L. Tinoko, TI Tikhonenko, EI Budovsky and others (1960 - 1974), the main features of the organization (packing) of the genetic material in bacteriophages became known.
In the late 1950s, the American scientist P. Doty found that when heated, DNA denaturation occurs, accompanied by the rupture of hydrogen bonds between base pairs and the divergence of complementary strands. This process has the character of a "spiral-coil" phase transition and resembles the melting of crystals. Therefore, Doty called the process of thermal denaturation of DNA DNA melting. Upon slow cooling, renaturation of the molecules occurs, that is, the reunification of the complementary halves.
The renaturation principle was used in 1960 by J. Marmur and K. Shildkraut to determine the degree of DNA "hybridizability" of different microorganisms. Subsequently E. Bolton and B. McCarthy improved this technique, proposing the method of so-called DNA-agar columns. This method turned out to be indispensable in studying the degree of homology of the nucleotide sequence of different DNA and elucidating genetic relationship different organisms. The open Doty DNA denaturation in combination with the chromatography described by J. Mandel and A. Hershey * (1960) on methylated albumin and density gradient centrifugation (the method was developed in 1957 by M. Meselson, F. Stahl, and D. Vinograd) is widely used for separation, isolation and analysis of individual complementary DNA strands For example, V. Shibalski (USA), using these techniques to separate the DNA of a lambda phage, showed in 1967 - 1969 that both phage strands are genetically active, and not one, as this was considered to be (S. Spigelman, 1961). It should be noted that the idea of \u200b\u200bthe genetic significance of both DNA strands of a lambda phage was first expressed in the USSR by S.E.Bresler (1961).
* (A. Hershey, together with M. Delbrück and S. Luria, were awarded the Nobel Prize in 1969 for their work on the genetics of bacteria and viruses.)
For understanding the organization and functional activity of the genome, the determination of the nucleotide sequence of DNA is of paramount importance. The search for methods for such a determination is carried out in many laboratories around the world. In the USA, M. Beer and his co-workers have been trying to establish the DNA sequence using electron microscopy since the late 1950s, but so far without success. In the early 50s, from the first works of Sinsheimer, Chargaff and other researchers on the enzymatic degradation of DNA, it became known that different nucleotides in the DNA molecule are distributed, albeit non-chaotically, but unevenly. According to the British chemist K. Barton (1961), pyrimidines (more than 70% of them) are concentrated mainly in the form of corresponding blocks. A. L. Mazin and B. F. Vanyushin (1968 - 1969) established that different DNAs have varying degrees of cohesion of pyrimidines and that in the DNA of animal organisms it noticeably increases with the transition from lower to higher. Thus, the evolution of organisms is reflected in the structure of their genomes. That is why, for understanding the evolutionary process as a whole, a comparative study of the structure of nucleic acids is of particular importance. Analysis of the structure of biologically important polymers and, first of all, DNA is extremely important for solving many particular problems of phylogenetics and taxonomy.
It is interesting to note that the English physiologist E. Lankester, who studied the hemoglobins of mollusks, and exactly 100 years ago anticipated the ideas of molecular biology, wrote: “Chemical differences between different species and genera of animals and plants are as important for clarifying the history of their origin as differences in If we could clearly establish the differences in the molecular organization and functioning of organisms, we would be able to understand much better the origin and evolution of different organisms than on the basis of morphological observations "*. The importance of biochemical studies for systematics was also emphasized by VL Komarov, who wrote that "all even purely morphological characters, on the basis of which we classify and establish species, are based on biochemical differences" **.
* (E. R. Lankester. Uber das Vorcommen von Haemoglobin in den Muskeln der Mollusken und die Verbreitung desselben in den lebendigen Organismen. - "Pfluger" s Archiv fur die gesammte Physiol., 1871, Bd 4, 319.)
** (V.L. Komarov. Selected works, vol. 1. M.-L., Publishing house of the Academy of Sciences of the USSR, 1945, p. 331.)
A. V. Blagoveshchensky and S. L. Ivanov took the first steps in our country in the 1920s to clarify some questions of the evolution and systematics of organisms on the basis of a comparative analysis of their biochemical composition (see Chapter 2). Comparative analysis of the structure of proteins and nucleic acids is now becoming an increasingly tangible aid for taxonomists (see Chapter 21). This method of molecular biology makes it possible not only to clarify the position of individual species in the system, but also makes us take a fresh look at the very principles of classification of organisms, and sometimes revise the entire system as a whole, as happened, for example, with the taxonomy of microorganisms. Undoubtedly, in the future, analysis of the genome structure will occupy a central place in the chemosystematics of organisms.
Deciphering the mechanisms of DNA replication and transcription was of great importance for the formation of molecular biology (see Chapter 24).
An important shift in solving the problem of protein biosynthesis is associated with advances in the study of nucleic acids. In 1941 T. Casperson (Sweden) and in 1942 J. Brachet (Belgium) drew attention to the fact that tissues with active protein synthesis contain an increased amount of RNA. They concluded that ribonucleic acids play a crucial role in protein synthesis. In 1953, E. Gale and D. Fox seemed to have obtained direct evidence of the direct participation of RNA in protein biosynthesis: according to their data, ribonuclease significantly suppressed the incorporation of amino acids into lysates of bacterial cells. Similar data were obtained by V. Alfrey, M. Delhi and A. Mirsky (1953) on liver homogenates. Later, E. Gale rejected the correct idea expressed by him about the leading role of RNA in protein synthesis, mistakenly believing that the activation of protein synthesis in the cell-free system occurred under the influence of some other substance of unknown nature. In 1954 P. Zamechnik, D. Littlefield, RB Khesin-Lurie and others found that the most active inclusion of amino acids occurs in RNA-rich fractions of subcellular particles - microsomes. P. Zamechnik and E. Keller (1953 - 1954) found that the incorporation of amino acids was markedly enhanced in the presence of the supernatant fraction under the conditions of ATP regeneration. P. Sikewitz (1952) and M. Hoagland (1956) isolated from the supernatant a protein fraction (pH 5 fraction), which was responsible for the sharp stimulation of the inclusion of amino acids in microsomes. Along with proteins, a special class of low-molecular-weight RNAs, which are now called transport RNAs (tRNAs), were found in the supernatant. In 1958, Hoagland and Zamechnik, as well as P. Berg, R. Sweet and F. Allen and many other researchers discovered that the activation of each amino acid requires its own special enzyme, ATP and specific tRNA. It became clear that tRNAs perform exclusively the function of adapters, that is, adaptations that find the place of the corresponding amino acid in the forming protein molecule on the nucleic acid matrix (mRNA). These studies fully confirmed the adapter hypothesis of F. Crick (1957), which provided for the existence of polynucleotide adapters in the cell, which are necessary for the correct arrangement of the amino acid residues of the synthesized protein on the nucleic matrix. Much later, the French scientist F. Chapville (1962) in the laboratory of F. Lipman (Nobel Prize, 1953) in the USA very wittily and unambiguously showed that the location of an amino acid in a synthesized protein molecule is completely determined by the specific tRNA to which it is attached. Crick's adapter hypothesis was developed by Hoagland and Zamechnik.
By 1958, the following main stages of protein synthesis became known: 1) activation of an amino acid by a specific enzyme from the "pH 5 fraction" in the presence of ATP with the formation of aminoacyladenylate; 2) attachment of an activated amino acid to a specific tRNA with the release of adenosine monophosphate (AMP); 3) binding of aminoacyl-tRNA (tRNA loaded with amino acid) with microsomes and incorporation of amino acids into the protein with the release of tRNA. Hoagland (1958) noted that guanosine triphosphate (GTP) is required at the last stage of protein synthesis.
After the discovery of tRNA, an active search for their fractionation and determination of the nucleotide sequence began. The greatest success was achieved by the American biochemist R. Holly. In 1965, he established the structure of alanine tRNA from yeast. Using ribonucleases (guanyl RNAse and pancreatic RNAse), Holly divided the nucleic acid molecule into several fragments, determined the nucleotide sequence in each of them separately, and then reconstructed the sequence of the entire alanine tRNA molecule. This way of analyzing the nucleotide sequence is called the block method. Holly's merit consisted mainly in the fact that he learned how to divide the RNA molecule not only into small pieces, as many had done before him, but also into large fragments (quarters and halves). This gave him the opportunity to correctly assemble separate small pieces together and thereby recreate the complete nucleotide sequence of the entire tRNA molecule (Nobel Prize, 1968).
This technique was immediately adopted by many laboratories around the world. Over the next two years, the primary structure of several tRNAs was deciphered in the USSR and abroad. A. A. Baev (1967) and colleagues for the first time established the sequence of nucleotides in yeast valine tRNA. To date, more than a dozen different individual tRNAs have been studied. A kind of record in determining the nucleotide sequence was set in Cambridge by F. Senger and G. Brownlee. These researchers developed a surprisingly elegant method for separating oligonucleotides and established the sequence of the so-called 5 S (ribosomal) RNA from E. coli cells (1968). This RNA consists of 120 nucleotide residues and, unlike tRNA, does not contain additional minor bases, which greatly facilitate the analysis of the nucleotide sequence, serving as unique landmarks for individual fragments of the molecule. At present, thanks to the use of the Sanger and Brownlee method, the work on the study of the sequence of long ribosomal RNAs and some viral RNAs in the laboratory of J. Ebel (France) and other researchers is progressing successfully.
AA Baev et al (1967) found that valine tRNA, cut in half, restores its macromolecular structure in solution and, despite a defect in the primary structure, has the functional activity of the original (native) molecule. This approach - reconstruction of a cut macromolecule after removal of certain fragments - has proven to be very promising. It is now widely used to elucidate the functional role of individual regions of certain tRNAs.
In recent years, great success has been achieved in the preparation of crystalline preparations of individual tRNAs. Nowadays, many tRNAs have already been crystallized in several laboratories in the USA and England. This made it possible to study the structure of tRNA using X-ray diffraction analysis. In 1970, R. Bock presented the first X-ray diffraction patterns and three-dimensional models of several tRNAs, created by him at the University of Wisconsin. These models help determine the localization of individual functionally active sites in tRNA and understand the basic principles of the functioning of these molecules.
Deciphering the nature of the genetic code (see Chapter 24), which, without exaggeration, can be regarded as the leading conquest of natural science in the 20th century, was of paramount importance for revealing the mechanism of protein synthesis and solving the problem of the specificity of this process.
R. Holly's disclosure of the primary structure of tRNA gave impetus to the works of G. Korana * (USA) on the synthesis of oligonucleotides and directed them to the path of synthesis of a certain biological structure - a DNA molecule encoding alanine tRNA. The first steps in the chemical synthesis of short oligonucleotides, made by the Koran almost 15 years ago, ended in 1970 with the first synthesis of a gene. The Korana and his collaborators first synthesized short fragments of 8-12 nucleotide residues from individual nucleotides by chemical means. These fragments with a given nucleotide sequence formed spontaneously double-stranded complementary pieces with an overlap of 4-5 nucleotides. Then these ready-made pieces in the desired order were alternately connected end-to-end using the enzyme DNA ligase. Thus, in contrast to the replication of DNA molecules, according to A. Kornberg ** (see Chapter 24), the Quran was able to re-create a natural double-stranded DNA molecule according to a pre-planned program in accordance with the tRNA sequence described by Holly. Similarly, work is now underway on the synthesis of other genes (MN Kolosov, Z. A. Shabarova, DG Knorre, 1970 - 1975).
* (G. Korana and M. Nirenberg were awarded the Nobel Prize in 1968 for the study of the genetic code.)
** (For the discovery of polymerase and DNA synthesis A. Kornberg, and for the synthesis of RNA S. Ochoa in 1959 was awarded the Nobel Prize.)
In the mid-1950s, it was believed that microsomes are the center of protein synthesis in the cell. The term microsomes was first introduced in 1949 by A. Claude to designate the fraction of small granules. Later it turned out that not the entire fraction of microsomes, consisting of membranes and granules, is responsible for protein synthesis, but only small ribonucleoprotein particles. In 1958, these particles were named ribosomes by R. Roberts.
Classical studies of bacterial ribosomes were carried out by A. Tissier and J. Watson in 1958 - 1959. Bacterial ribosomes were found to be somewhat smaller than plants and animals. J. Littleton (1960), M. Clarke (1964), and E.N.Svetailo (1966) showed that the ribosomes of chloroplasts of higher plants and mitochondria belong to the bacterial type. A. Tissier and others (1958) found that ribosomes dissociate into two unequal subunits containing one RNA molecule. In the late 1950s, it was believed that each molecule of ribosomal RNA consists of several short fragments. However, A.S.Spirin in 1960 showed for the first time that RNA in subparticles is represented by a continuous molecule. D. Waller (1960), having separated ribosomal proteins by electrophoresis in starch gel, found that they are very heterogeneous. At first, many doubted Waller's data, since it seemed that the ribosome protein should be strictly homogeneous, such as the TMV protein. At present, as a result of studies by D. Waller, R. Trout, P. Traub and other biochemists, it has become known that the composition of the ribosomal particles proper includes more than 50 proteins that are completely different in structure. A.S. Spirin in 1963 was the first to unroll ribosomal subunits and show that ribosomes are a compactly twisted ribonucleoprotein strand that can unfold under certain conditions. 1967-1968 M. Nomura completely reconstructed a biologically active subunit from ribosomal RNA and protein and even obtained such ribosomes in which the protein and RNA belonged to different microorganisms.
Until now, the role of ribosomal RNA is unclear. It is assumed that it is that unique specific matrix on which, during the formation of a ribosomal particle, each of the numerous ribosomal proteins finds a strictly defined place (A.S. Spirin, 1968).
A. Rich (1962) discovered aggregates of several ribosomes interconnected by a mRNA strand. These complexes were called polysomes. The discovery of polysomes allowed Rich and Watson (1963) to suggest that the synthesis of the polypeptide chain occurs on the ribosome, which, as it were, moves along the mRNA chain. As the ribosome moves along the mRNA chain, information is read in the particle and the polypeptide chain of the protein is formed, and new ribosomes alternately attach to the released read end of the mRNA. From the data of Rich and Watson it followed that the significance of the polysome in the cell consists in the mass production of protein by sequential reading of the matrix by several ribosomes at once.
As a result of research by M. Nirenberg, S. Ochoa, F. Lipman, G. Korana and others in 1963 - 1970. it became known that along with mRNA, ribosomes, ATP and aminoacyl-tRNA, a large number of various factors are involved in the translation process, and the translation process itself can be conditionally divided into three stages - initiation, translation itself, and termination.
Translation initiation means the synthesis of the first peptide bond in the ribosome - template polynucleotide - aminoacyl-tRNA complex. This initiator activity is not possessed by any aminoacyl-tRNA, but formylmethionyl-tRNA. This substance was first isolated in 1964 by F. Senger and K. Marker. S. Bretcher and K. Marker (1966) showed that the initiator function of formylmethionyl-tRNA is due to its increased affinity for the peptidyl center of the ribosome. For the beginning of translation, some protein initiation factors, which were isolated in the laboratories of S. Ochoa, F. Gro and other research centers, are also extremely important. After the formation of the first peptide bond in the ribosome, translation itself begins, i.e., the sequential attachment of the aminoacyl residue to the C-terminus of the polypeptide. Many details of the broadcasting process were studied by K. Monroe and J. Bishop (England), I. Rykhlik and F. Schorm (Czechoslovakia), F. Lipman, M. Bretcher, W. Gilbert (USA) and other researchers. In 1968 A.S.Spirin proposed an original hypothesis to explain the mechanism of the ribosome. The driving mechanism providing all the spatial movements of tRNA and mRNA during translation is the periodic opening and closing of ribosome subunits. The end of translation is encoded in the readable matrix itself, which contains the termination codons. As shown by S. Brenner (1965 - 1967), such codons are the triplets UAA, UAG, and UGA. M. Capecchi (1967) also identified special protein termination factors. AS Spirin and LP Gavrilova described the so-called "non-enzymatic" protein synthesis in ribosomes (1972 - 1975) without the participation of protein factors. This discovery is important for understanding the origin and evolution of protein biosynthesis.
After the problem of the specificity of protein synthesis, the problem of regulation of protein synthesis, or, which is the same thing, regulation of gene activity, turned out to be in the first place in molecular biology.
The functional inequality of cells and the associated repression and activation of genes have long attracted the attention of geneticists, but until recently the real mechanism of control of gene activity remained unknown.
The first attempts to explain the regulatory activity of genes were associated with the study of histone proteins. Even the Steadman spouses * in the early 40s of the XX century. expressed the idea that it is histones that can play the main role in this phenomenon. Subsequently, they received the first clear data on the differences in the chemical nature of histone proteins. At present, the number of facts supporting this hypothesis is increasing every year.
* (E. Stedman, E. Stedman. The basic proteins of cell nuclei - Phylosoph. Trans. Roy. Soc. London, 1951, v. 235, 565 - 595.)
At the same time, more and more data are accumulating indicating that the regulation of gene activity is a much more complex process than the simple interaction of gene regions with molecules of histone proteins. In 1960-1962 In the laboratory of RB Khesin-Lurie, it was found that phage genes begin to be read non-simultaneously: the T2 phage genes can be divided into early genes, the functioning of which occurred in the first minutes of infection of the bacterial cell, and late genes, which began to synthesize mRNA after the completion of the early genes.
In 1961, the French biochemists F. Jacob and J. Monod proposed a scheme for the regulation of gene activity, which played an exceptional role in understanding the regulatory mechanisms of the cell in general. According to Jacob and Monod's scheme, in addition to structural (informational) genes, DNA contains also regulator genes and operator genes. The regulator gene encodes the synthesis of a specific substance, a repressor, which can be attached to both the inducer and the operator gene. The operator gene is linked to structural genes, and the regulator gene is located at some distance from them. If there is no inductor in the environment, for example, lactose, then the repressor synthesized by the regulator gene binds to the operator gene and, blocking it, turns off the work of the entire operon (the block of structural genes together with the operator that controls them). The enzyme does not form under these conditions. If an inducer (lactose) appears in the medium, then the product of the regulator gene - the repressor - binds to lactose and removes the block from the operator gene. In this case, the work of the structural gene that encodes the synthesis of the enzyme becomes possible, and the enzyme (lactose) appears in the medium.
According to Jacob and Monod, this regulation scheme is applicable to all adaptive enzymes and can take place both during repression, when enzyme formation is suppressed by an excess of the reaction product, and during induction, when the addition of a substrate causes enzyme synthesis. In 1965, Jacob and Monod were awarded the Nobel Prize for their studies on the regulation of gene activity.
Initially, this scheme seemed too far-fetched. However, it later became clear that gene regulation according to this principle takes place not only in bacteria, but also in other organisms.
Since 1960, a prominent place in molecular biology has been occupied by studies of genome organization and chromatin structure in eukaryotic organisms (J. Bonner, R. Britten, W. Alfrey, P. Walker, Yu.S. Chentsov, I.B. Zbarsky, etc. .) and the regulation of transcription (A. Mirsky, G. P. Georgiev, M. Bernstil, D. Goll, R. Tsanev, R. I. Salganik). The nature of the repressor remained unknown and controversial for a long time. In 1968 M. Ptashne (USA) showed that a protein is a repressor. He isolated it in the laboratory of J. Watson and found that the repressor does indeed have an affinity for the inducer (lactose) and at the same time "recognizes" the gene operator of the lac operon and binds specifically to it.
In the last 5-7 years, data have been obtained on the presence of another control cell of gene activity - a promoter. It turned out that in the vicinity of the operator site, to which the product synthesized on the gene-regulator - the protein substance of the repressor, is attached, there is another site, which should also be attributed to the members of the regulatory system of gene activity. A protein molecule of the RNA polymerase enzyme is attached to this site. In the promoter region, mutual recognition of the unique nucleotide sequence in DNA and the specific configuration of the RNA polymerase protein should occur. The implementation of the process of reading genetic information with a given gene sequence of the operon adjacent to the promoter will depend on the recognition efficiency.
In addition to the scheme described by Jacob and Monod, there are other mechanisms of gene regulation in the cell. F. Jacob and S. Brenner (1963) found that the regulation of bacterial DNA replication is controlled in a certain way by the cell membrane. The experiments of Jacob (1954) on the induction of various prophages convincingly showed that, under the influence of various mutagenic factors, selective replication of the prophage gene begins in the cell of lysogenic bacteria, and replication of the host genome is blocked. In 1970, F. Bell reported that small DNA molecules can enter the cytoplasm from the nucleus and be transcribed there.
Thus, the regulation of gene activity can be carried out at the level of replication, transcription, and translation.
Significant advances have been made in studying the regulation of not only enzyme synthesis, but also their activity. The phenomena of regulation of the activity of enzymes in the cell were pointed out back in the 1950s by A. Novik and L. Szilard. G. Umbarger (1956) established that in the cell there is a very rational way of suppressing the enzyme activity by the final product of the feedback chain of reactions. As was established by J. Monod, J. Changer, F. Jacob, A. Purdy and other researchers (1956 - 1960), the regulation of enzyme activity can be carried out according to the allosteric principle. An enzyme or one of its subunits, in addition to an affinity for a substrate, has an affinity for one of the products of the chain of reactions. Under the influence of such a signal product, the enzyme changes its conformation so that it loses its activity. As a result, the entire chain of enzymatic reactions is turned off at the very beginning. D. Weeman and R. Woodward (1952; Nobel Prize winner, 1965) pointed out the essential role of protein conformational changes in enzymatic reactions, and in a certain sense, the presence of an allosteric effect.
As a result of the works of T. Osborne, G. Hofmeister, A. Gürber, F. Schulz and many others at the end of the 19th century. many animal and plant proteins were obtained in crystalline form. At about the same time, with the help of various physical methods the molecular weights of some proteins have been established. Thus, in 1891 A. Sabaneev and N. Aleksandrov reported that the molecular weight of ovalbumin was 14,000; in 1905, E. Reid established that the molecular weight of hemoglobin is 48,000. The polymeric structure of proteins was discovered in 1871 by G. Glazivets and D. Haberman. The idea of \u200b\u200bthe peptide bond of individual amino acid residues in proteins was put forward by T. Curtius (1883). Work on the chemical condensation of amino acids (E. Schaal, 1871; G. Schiff, 1897; L. Balbiano and D. Truschiatti, 1900) and the synthesis of heteropolypeptides (E. Fischer, 1902-1907, Nobel Prize, 1902) led to the development of basic principles chemical structure of proteins.
The first crystalline enzyme (urease) was obtained in 1926 by J. Sumner (Nobel Prize, 1946), and in 1930, J. Northrop (Nobel Prize, 1946) received crystalline pepsin. After these works, it became clear that the enzymes are of a protein nature. In 1940 M. Kunits isolated crystalline RNAse. By 1958, more than 100 crystalline enzymes and over 500 enzymes isolated in non-crystalline form were already known. Obtaining highly purified preparations of individual proteins contributed to the deciphering of their primary structure and macromolecular organization.
Of great importance for the development of molecular biology in general and human genetics, in particular, was the discovery by L. Pauling (1940) of abnormal hemoglobin S, isolated from the erythrocytes of people with a severe hereditary disease - sickle cell anemia. In 1955 - 1957 V. Ingram used the method of "fingerprints" developed by F. Senger (spots formed by individual peptides during chromatography on paper) to analyze the products of hemoglobin S hydrolysis with alkali and trypsin. In 1961, Ingram reported that hemoglobin S differs from normal hemoglobin only in the nature of one amino acid residue: in normal hemoglobin in the seventh position of the chain there is a glutamic acid residue, and in hemoglobin S there is a valine residue. This fully confirmed (1949) Pauling's assumption that sickle cell anemia is a disease of molecular nature. An inherited change in only one amino acid residue in each half of the hemoglobin macromolecule leads to the fact that hemoglobin loses its ability to easily dissolve at low oxygen concentrations and begins to crystallize, which leads to a violation of the cell structure. These studies have clearly shown that the structure of a protein is a strictly defined amino acid sequence that is encoded in the genome. The exceptional importance of the primary structure of a protein in the formation of a unique biologically active conformation of a macromolecule was evidenced by the works of K. Anfinsen (1951). Anfinsen showed that the biologically active macrostructure of pancreatic ribonuclease, lost as a result of reduction, is predetermined by the amino acid sequence and can reappear spontaneously upon oxidation of SH-groups of cysteine \u200b\u200bresidues with the formation of disulfide crosslinks at strictly defined sites of the enzyme peptide chain.
To date, the mechanism of action of a large number of enzymes has been studied in detail and the structure of many proteins has been determined.
In 1953 F. Senger established the amino acid sequence of insulin. : This protein consists of two polypeptide chains linked by two disulfide cross-links. One of the chains contains only 21 amino acid residues, while the other contains 30 residues. Sanger spent about 10 years to decipher the structure of this relatively simple protein. In 1958, he was awarded the Nobel Prize for this outstanding research. After the creation of an automatic amino acid analyzer by W. Stein and S. Moore (1957), the identification of the products of partial hydrolysis of proteins was significantly accelerated. In 1960 Stein and Moore already reported this. that they were able to determine the sequence of ribonuclease, the peptide chain of which is represented by 124 amino acid residues. In the same year in the laboratory of G. Schramm in Tübingen (Germany) F. Anderer and others determined the amino acid sequence in the TMV protein. Then the amino acid sequence was determined in myoglobin (A. Edmunson) and α- and β-chains of human hemoglobin (G. Braunitzer, E. Schroeder and others), lysozyme from chicken egg protein (J. Jollet, D. Keyfield). In 1963 F. Schorm and B. Keil (Czechoslovakia) established the sequence of amino acids in the chymotrypsinogen molecule. In the same year, the amino acid sequence of trypsinogen was determined (F. Schorm, D. Walsh). In 1965, K. Takahashi established the primary structure of ribonuclease T1. Then the amino acid sequence was determined in several more proteins.
As you know, the final proof of the correctness of the definition of a particular structure is its synthesis. In 1969 R. Merifield (USA) was the first to chemically synthesize pancreatic ribonuclease. Using his method of synthesis on a solid-phase carrier, Maryfield added one amino acid after another to the chain in accordance with the sequence described by Stein and Moore. As a result, he obtained a protein that was identical in its qualities to pancreatic ribonuclease A. For the disclosure of the structure of ribonuclease V. Stein, S. Moore and K. Anfinsen were awarded the Nobel Prize in 1972. This natural protein synthesis opens up great prospects, indicating the possibility of creating any proteins in accordance with a pre-planned sequence.
From the X-ray diffraction studies of W. Astbury (1933) it followed that the peptide chains of protein molecules are twisted or folded in some strictly defined way. Since that time, many authors have expressed various hypotheses about the methods of folding protein chains, but until 1951 all models remained speculative constructions that did not correspond to experimental data. In 1951 L. Pauling and R. Corey published a series of brilliant works in which the theory of the secondary structure of proteins - the theory of the α-helix - was finally formulated. Along with this, it became known that proteins also have a tertiary structure: the α-helix of the peptide chain can be folded in a certain way, forming a rather compact structure.
In 1957, J. Kendrew and his colleagues first proposed a three-dimensional model of the structure of myoglobin. This model was then refined over several years, until in 1961 the final work with the characterization of the spatial structure of this protein appeared. In 1959 M. Perutz and colleagues established the three-dimensional structure of hemoglobin. The researchers spent more than 20 years on this work (the first radiographs of hemoglobin were obtained by Perutz in 1937). Since the hemoglobin molecule consists of four subunits, then, having deciphered its organization, Perutz thereby first described the quaternary structure of a protein. Kendrew and Perutz were awarded the Nobel Prize in 1962 for their work on the determination of the three-dimensional structure of proteins.
The creation of a spatial model of the hemoglobin structure by Perutz ENABLED. come closer to understanding the mechanism of the functioning of this protein, which, as you know, carries out the transfer of oxygen in animal cells. Back in 1937, F. Gaurowitz came to the conclusion that the interaction of hemoglobin with oxygen, air should be accompanied by a change in the protein structure. In the 1960s, Perutz and his collaborators discovered a noticeable shift in hemoglobin chains after oxidation, caused by a shift in iron atoms as a result of binding with oxygen. On this basis, ideas about the "respiration" of protein macromolecules were formed.
In 1960, D. Phillips and his colleagues began X-ray structural studies of the lysozyme molecule. By 1967, they more or less accurately succeeded in establishing the details of the organization of this protein and the localization of individual atoms in its molecule. In addition, Phillips found out the nature of the addition of lysozyme to the substrate (triacetylglucosamine). This made it possible to recreate the mechanism of this enzyme. Thus, knowledge of the primary structure and macromolecular organization made it possible not only to establish the nature of the active centers of many enzymes, but also to fully reveal the mechanism of functioning of these macromolecules.
The use of electron microscopy methods helped to reveal the principles of the macromolecular organization of such complex protein formations as collagen, fibrinogen threads, contractile muscle fibrils, etc. At the end of the 50s, models of the muscular contractile apparatus were proposed. The discovery of the ATPase activity of myosin by V.A.Engel'gardt and M.N.Lyubimova (1939) was of exceptional importance for understanding the mechanism of muscle contraction. This meant that the act of muscle contraction is based on a change in the physicochemical properties and macromolecular organization of the contractile protein under the influence of adenosine triphosphoric acid (see also Chapter 11).
Virological studies were essential to understanding the principles of assembly of biological structures (see Chapter 25).
Major advances in modern molecular biology have been achieved mainly as a result of the study of nucleic acids. Nevertheless, even in this area, far from all problems have been resolved. In particular, decoding of the entire nucleotide sequence of the genome will require great efforts. This problem, in turn, is inextricably linked with the problem of DNA heterogeneity and requires the development of new advanced methods of fractionation and isolation of individual molecules from the total genetic material of the cell.
So far, efforts have mainly focused on the separate study of proteins and nucleic acids. In the cell, these biopolymers are inextricably linked with each other and function mainly in the form of nucleoproteins. Therefore, the need to study the interaction of proteins and nucleic acids has now become especially acute. The problem of recognition of certain regions of nucleic acids by proteins is highlighted. Steps have already been outlined towards the study of such an interaction of these biopolymers, without which a complete understanding of the structure and functions of chromosomes, ribosomes and other structures is inconceivable. Without this, it is also impossible to understand the regulation of gene activity and finally decipher the principles of the protein-synthesizing mechanisms. After the work of Jacob and Monod, some new data appeared on the regulatory role of membranes in the synthesis of nuclear material. This poses the problem of a deeper study of the role of membranes in the regulation of DNA replication. In general, the problem of regulating gene activity and cellular activity in general has become one of the most important problems of modern molecular biology.
Biophysics developed in close connection with the problems of molecular biology. Interest in this area of \u200b\u200bbiology was stimulated, on the one hand, by the need for a comprehensive study of the effects of various kinds of radiation on the body, on the other, by the need to study the physical and physicochemical foundations of life phenomena occurring at the molecular level.
Acquisition of accurate information about molecular structures and processes occurring in them has become possible as a result of the use of new fine physicochemical methods. On the basis of the achievements of electrochemistry, it was possible to improve the method of measuring bioelectric potentials by using ion-selective electrodes (G. Eisenman, B. P. Nikol'skii, Khuri, 50s - 60s). Infrared spectroscopy (with the use of laser devices), which makes it possible to investigate conformational changes in proteins, is becoming increasingly common (I. Plotnikov, 1940). Valuable information is also provided by the method of electron paramagnetic resonance (EK Zavoisky, 1944) and the biochemoluminescent method (BN Tarusov et al., 1960), which, in particular, make it possible to judge the transport of electrons during oxidative processes.
By the 50s, biophysics is already gaining a strong position. There is a need for the training of qualified specialists. If in 1911 in Europe only at the University of Pecs, in Hungary, there was a department of biophysics, then by 1973 such departments exist in almost all large universities.
In 1960, the International Society of Biophysicists was organized. In August 1961, the first International Biophysical Congress was held in Stockholm. The second congress was held in 1965 in Paris, the third in 1969 in Boston, and the fourth in 1972 in Moscow.
In biophysics, there is a clear distinction between two areas of different content - molecular biophysics and cellular biophysics. This distinction also receives organizational expression: separate departments of these two areas of biophysics are created. At Moscow University, the first department of biophysics was created in 1953 at the Faculty of Biology and Soil Science; a little later, the Department of Biophysics was established at the Faculty of Physics. Departments in many other universities were organized on the same principle.
In recent years, the link between molecular biophysics and molecular biology has been increasingly strengthened, and now it is sometimes difficult to determine where the interface between them lies. In a general attack on the problem of hereditary information, such cooperation of biophysics with molecular biology is inevitable.
The main direction of research is the study of the physics of nucleic acids - DNA and RNA. The use of the above methods and, first of all, X-ray structural analysis contributed to the deciphering of the molecular structure of nucleic acids. At present, intensive research is underway to study the behavior of these acids in solutions. At the same time, special attention is paid to the conformational transitions "helix-coil", studied by changes in viscosity, optical and electrical indicators. In connection with the study of the mechanisms of mutagenesis, research is being developed to study the effect of ionizing radiation on the behavior of nucleic acids in solutions, as well as the effect of radiation on nucleic acids of viruses and phages. The influence of ultraviolet radiation was subjected to a comprehensive analysis, some spectral regions of which are known to be well absorbed by nucleic acids. Large specific gravity in this kind of research is the detection of active radicals of nucleic acids and proteins by the method of electron paramagnetic resonance. The use of this method is associated with the emergence of a whole independent direction.
The problem of coding DNA and RNA information and its transfer during protein synthesis has long been of interest to molecular biophysics, and physicists have repeatedly expressed certain considerations on this matter (E. Schrödinger, G. Gamow). Deciphering the genetic code has caused numerous theoretical and experimental studies on the structure of the DNA helix, the mechanism of sliding and twisting of its threads, on the study of the physical forces involved in these processes.
Molecular biophysics provides molecular biology with significant assistance in studying the structure of protein molecules using X-ray diffraction analysis, which was first used in 1930 by J. Bernal. It was as a result of the use of physical methods in combination with biochemical (enzymatic methods) that the molecular conformation and sequence of amino acids in a number of proteins were revealed.
Modern electron microscopic studies, revealing the presence of complex membrane systems in cells and its organelles, stimulated attempts to understand their molecular structure (see Chapters 10 and 11). Studied in vivo chemical composition membranes and, in particular, the properties of their lipids. It was found that the latter are capable of peroxidation and non-enzymatic reactions of chain oxidation (Yu.A. Vladimirov and F.F. Litvin, 1959; B.N.Tarusov et al., 1960; I.I. Ivanov, 1967), leading to violation of membrane functions. Methods of mathematical modeling were also used to study the composition of membranes (V. Ts. Presman, 1964 - 1968; M. M. Shemyakin, 1967; Yu. A. Ovchinnikov, 1972).
A significant event in the history of biophysics was the formation in the 50s of a clear understanding of the thermodynamics of biological processes, as a result of which the assumptions about the possibility of independent generation of energy in living cells in spite of the second law of thermodynamics have finally disappeared. Understanding the operation of this law in biological systems connected with the introduction by the Belgian scientist I. Prigozhin (1945) * into biological thermodynamics of the concept of open systems exchanging energy and matter with the external environment. Prigogine showed that positive entropy is formed in living cells during working processes in accordance with the second law of thermodynamics. The equations he introduced determined the conditions under which a so-called stationary state arises (previously it was also called dynamic equilibrium), in which the amount of free energy (negentropy) entering the cells with food compensates for its consumption, and positive entropy is derived. This discovery has supported the general biological idea of \u200b\u200ban inextricable connection between the external and internal environment of cells. It marked the beginning of the real study of the thermodynamics of "alive" systems, including the modeling method (A. Burton, 1939; A. G. Pasynsky, 1967).
* (The general theory of open systems was first put forward by L. Bertalanffy in 1932.)
According to the basic principle of biothermodynamics, a necessary condition for the existence of life is stationarity in the development of its biochemical processes, for the implementation of which the coordination of the rates of numerous metabolic reactions is necessary. On the basis of new biophysical thermodynamics, a direction has emerged that distinguishes external and internal factors that ensure this coordination of reactions and make it stable. Over the past two decades, a large role in maintaining a steady state of the system of inhibitors and especially antioxidants has been revealed (BN Tarusov and AI Zhuravlev, 1954, 1958). It has been established that the reliability of stationary development is associated with environmental factors (temperature) and physicochemical properties of the cell environment.
Modern principles of biothermodynamics have made it possible to give a physicochemical interpretation of the adaptation mechanism. According to our data, adaptation to environmental conditions can occur only if, when changing them, the body is able to establish stationarity in the development of biochemical reactions (BN Tarusov, 1974). The question arose about the development of new methods that would make it possible to assess the steady state in vivo and predict its possible violations. The introduction of the cybernetic principles of self-regulating systems into biothermodynamics and research into the processes of biological adaptation promises great benefits. It became clear that in order to resolve the issue of the stability of a steady state, it is important to take into account the so-called disturbing factors, which include, in particular, non-enzymatic reactions of lipid oxidation. In recent years, more and more studies of the processes of peroxidation in the lipid phases of living cells and the growth of active radical products that disrupt the regulatory functions of membranes have been expanding. The source of information about these processes is both the detection of active peroxide radicals and peroxide compounds of biolipids (A. Tappel, 1965; I. I. Ivanov, 1965; EB Burlakova, 1967 and others). To detect radicals, biochemoluminescence is used, which occurs in the lipids of living cells during their recombination.
On the basis of physicochemical ideas about the stability of a stationary state, biophysical ideas arose about the adaptation of plants to changes in environmental conditions as a violation of inhibiting antioxidant systems (B.N. Tarusov, Ya.E. Doskoch, B.M. Kitlaev, A.M. , 1968 - 1972). This opened up the possibility of evaluating properties such as frost resistance and salt tolerance, as well as making appropriate predictions when breeding agricultural plants.
In the 50s, a superweak glow was discovered - biochemoluminescence of a number of biological objects in the visible and infrared parts of the spectrum (BN Tarusov, AI Zhuravlev, AI Polivoda). This became possible as a result of the development of methods for recording superweak light fluxes using photomultiplier tubes (L.A. Kubetsky, 1934). As a result of biochemical reactions occurring in a living cell, biochemoluminescence makes it possible to judge important oxidative processes in the electron transport chains between enzymes. The discovery and study of biochemoluminescence is of great theoretical and practical importance. Thus, BN Tarusov and Yu. B. Kudryashov note the important role of the oxidation products of unsaturated fatty acids in the mechanism of the onset of pathological conditions developing under the influence of ionizing radiation during carcinogenesis and other disorders of normal cell functions.
In the 1950s, in connection with the rapid development of nuclear physics, radiobiology emerged from biophysics, which studies the biological effect of ionizing radiation. The production of artificial radioactive isotopes, the creation of thermonuclear weapons, atomic reactors and the development of other forms of practical use of atomic energy have posed with all their urgency the problem of protecting organisms from the harmful effects of ionizing radiation, the development of theoretical foundations prevention and treatment of radiation sickness. To do this, it was necessary, first of all, to find out which components of the cell and metabolic links are most vulnerable.
The object of the study of biophysics and radiobiology was the elucidation of the nature of the primary chemical reactions that occur in living substrates under the influence of radiation energy. Here it was important not only to understand the mechanisms of this phenomenon, but also to be able to influence the process of exchanging physical energy for chemical energy, to reduce its coefficient of "useful" action. Work in this direction laid the foundation for the study of the school of N. N. Semenov (1933) in the USSR and D. Hinshelwood (1935) in England.
An important place in radiobiological research has been occupied by the study of the degree of radiation resistance of various organisms. It was found that increased radioresistance (for example, of rodents of deserts) is due to the high antioxidant activity of lipids of cell membranes (M. Chang et al., 1964; NK Ogryzov et al., 1969). It turned out that tocopherols, vitamin K and thio compounds play an important role in the formation of the antioxidative properties of these systems (II Ivanov et al., 1972). In recent years, studies of the mechanisms of mutagenesis have also attracted great attention. For this purpose, the effect of ionizing radiation on the behavior of nucleic acids and proteins in vitro, as well as in viruses and phages, is being studied (A. Gustafson, 1945-1950).
The struggle for a further increase in the effectiveness of chemical protection, the search for more effective inhibitors and principles of inhibition remain the main tasks of biophysics in this direction.
Research into the excited states of biopolymers, which determine their high chemical activity, has advanced. The most successful was the study of excited states arising at the primary stage of photobiological processes - photosynthesis and vision.
Thus, a substantial contribution has been made to the understanding of the primary activation of molecules of plant pigment systems. It has been established that the transfer (migration) of the energy of excited states without losses from the activated pigments to other substrates is of great importance. The theoretical works of A. N. Terenin (1947 and later) played an important role in the development of these concepts. AA Krasnovsky (1949) discovered and investigated the reaction of reversible photochemical reduction of chlorophyll and its analogs. There is now a general belief that in the near future it will be possible to reproduce photosynthesis in artificial conditions (see also chapter 5).
Biophysicists continue to work to uncover the nature of muscle contraction and the mechanisms of nerve excitation and conduction (see Chapter 11). Studies of the mechanisms of the transition from an excited state to a normal state have also acquired topical importance. An excited state is now considered as a result of an autocatalytic reaction, and inhibition is a consequence of a sharp mobilization of inhibitory antioxidant activity as a result of molecular rearrangements in compounds such as tocopherol (I.I. Ivanov, O.R. Kols, 1966; O.R. Kols, 1970).
The most important general problem of biophysics remains the cognition of the qualitative physicochemical characteristics of living matter. Properties such as the ability of living biopolymers to selectively bind potassium or to polarize electric current cannot be preserved even with the most careful extraction from the body. Therefore, cell biophysics continues to intensively develop criteria and methods for the in vivo study of living matter.
Despite the youth of molecular biology, the successes it has achieved in this area are truly stunning. In a relatively short time, the nature of the gene and the basic principles of its organization, reproduction and functioning have been established. Moreover, not only the multiplication of genes in vitro has been carried out, but for the first time the complete synthesis of the gene itself has been completed. The genetic code has been fully deciphered and the most important biological problem of the specificity of protein biosynthesis has been resolved. The main pathways and mechanisms of protein formation in the cell have been identified and investigated. The primary structure of many transport RNAs - specific adapter molecules that translate the language of nucleic acid templates into the language of the amino acid sequence of the synthesized protein - has been fully determined. The amino acid sequence of many proteins has been fully deciphered and the spatial structure of some of them has been established. This made it possible to clarify the principle and details of the functioning of enzyme molecules. Chemical synthesis of one of the enzymes, ribonuclease, has been carried out. The basic principles of the organization of various subcellular particles, many viruses and phages have been established, and the main pathways of their biogenesis in the cell have been unraveled. Approaches to understanding the ways of regulating gene activity and elucidating the regulatory mechanisms of vital activity have been revealed. Already a simple list of these discoveries indicates that the second half of the XX century. was marked by tremendous progress in biology, which is primarily due to the in-depth study of the structure and functions of biologically important macromolecules - nucleic acids and proteins.
The achievements of molecular biology are already being used in practice today and are bearing tangible results in medicine, agriculture and some industries. There is no doubt that the impact of this science will increase every day. However, the main result should still be considered that, under the influence of the successes of molecular biology, confidence in the existence of unlimited possibilities for unlocking the most intimate secrets of life has been strengthened.
In the future, apparently, new ways of studying the biological form of the motion of matter will be discovered - biology will move from the molecular level to the atomic level. However, now there is, perhaps, not a single researcher who could reasonably predict the development of molecular biology even for the next 20 years.
The tarot cards are the most famous and most mysterious of all known ...

07/09/2015 Fortune-telling on Tarot cards is a kind of "aerobatics" in ...

The word runa means "secret" and goes back to the oldest root meaning ...